ACADEMY & GALLERY
1998, 2007 by L. C. Chin. Work in progress. So genetics, historical linguistics, and most of the archaeological evidences have all converged to convince us that we -- no matter what our color and cultural heritage -- are all brothers and sisters having descended from the same parents. Now we can go further. Since we can no longer subscribe to any notion of an independently existing soul -- a soul that leaves our body upon death and comes back to reincarnate in other human and animal bodies: Seelenwanderung or metempsychosis, as the ancient Greeks and Hindus believed -- we cannot now base our refrain from cruelty toward and consumption of animals, as Pythagoras was once said to have done, on the thinking that all life -- from gods through humans to animals -- is intimately linked because "the same spirit (soul) runs through everything in the cosmos" (πνευμα το δια παντος του κόσμου διηκον). But now another discovery has furnished a foundation for the old Pythagorean doctrine of the kinship of all living beings (die Lehre von der Verwandtschaft der Lebewesen), for our spiritual sentiment that all life is intimately linked -- we have now a new tool for the dismantling of the illusion of our separation from all the wonderful life around us -- and that is evolution, the teaching that all life on earth is descended from the same ancestor. We say now, "The same ancestry runs through every living being on earth." Indeed, we have already remarked on the spiritual implication of the theory of evolution: that we are brothers and sisters with all the animals we both domesticate as pets and breed and torture as food, with all the insects we find beautiful and abhor, with all the trees and plant life we ignore or exploit, and even with all the bacteria that we find harmful and try to ward off. Life is One because it all once came from the One. To feel the connectedness of all life through the study of evolution implies reading all those seemingly boring college textbooks on evolution in a new light; it means using them as tools for the dismantling of our illusion of separation, not just reading them in a disinterested academic attitude in order to pass a test or prepare for research; that is to say, we must regard them as religious text, even, and get ecstatic from their reading: books that we use here such as Colbert and Morales, Evolution of the Vertebrates, 4th ed.; Lynn Margulis et al, Five Kingdoms; Scagel, Bandoni, Maze, Rouse, Schofield, Stein, Plants: An Evolutionary Survey, and sections on evolution in Campbell, Mitchell, Reece, Biology, 2nd ed. You should get ecstactic from reading these books because you are learning about your ancestry, your heritage, your past, and your family connection with all the living beings around you. Even when you look at more specialized books (such as Silvia J. and Stephen A. Czerkas, Dinosaurs: A Global View) or websites (such as Frank Muster's: www.leute.server.de/frankmuster) on dinosaurs you should try to think that you are opening up the pages of your genealogy books. So we shall continue the anamnesis of our ancestry beyond the human realm by tracing the lineage of our ancestry backward, selecting out the more significant ones of our ancestors -- because we have only a limited amount of time here -- and visualizing and meditating upon them. In the following when you come to each of your ancestors so selected, you should realize that that is you, that you were once that. Try to imagine yourself being transformed into that ancestor of yours -- that was your former form -- and think of yourself as going through a series of transformation as your ancestral lineage passes before your eyes: evolution is how you have evolved. As we trace our ancestry backward in time, the first of our ancestors that we come to is Australopithecus afarensis, the so-called "Lucy", who lived about 3.5 million years ago. She stood no more than 120 centimeters. "Lucy" was the earliest species we know of that was on our lineage after our lineage separated from that leading to chimpanzees (Pan), and was the ancestor we have in common with all those cousins of ours that have gone extinct: Next we come to 55 million years ago. What does our ancestor look like at this time? Our ancestor at this time is a Plesiadapid, the common ancestor we share with all other primates. We must now imagine ourselves transforming into a Tupaia-like creature. Tupaia today resembles most the ancestor of all primates. "Morphologically the tree shrews [Tupaia] are so close to the line of demarcation between insectivores and primates that even if they are included in the former mammalian order, they, nevertheless, give us some insight into the adaptations that were leading from primitive eutherian [i.e. placental] mammals toward the first primates" (Colbert and Morales, ibid., p. 274).
The evolutionary origin of primates within the mammalian lineages (Class Mammalia) Primates constitute one of the twenty or so lineages (Order) within the infra class Eutheria Colbert and Morales, ibid., p. 242. Next stop in the tracing of our ancestry is the mammal-like reptile (säugetierähnliche Reptil) eotitanosuchus, an early gorgonopsian, which is ancestral to cynodonts, which in turn give rise to ictidosaurus (e.g. Diarthrognathus, closest to mammals). Eotitanosuchus is at the root of divergence between theriodonts and anomodonts (dinocephalians and dicynodonts), both of which belong within Therapsida, which originated from pelycosaurs, the early synapsid. (Synapsida (mammal-like reptile) is made up of both the pelycosaurs and therapsids.) Eotitanosuchus is a primitive carnivore, "primitive" in the sense that its small temple opening behind its eyes allowed only for a weak bite. 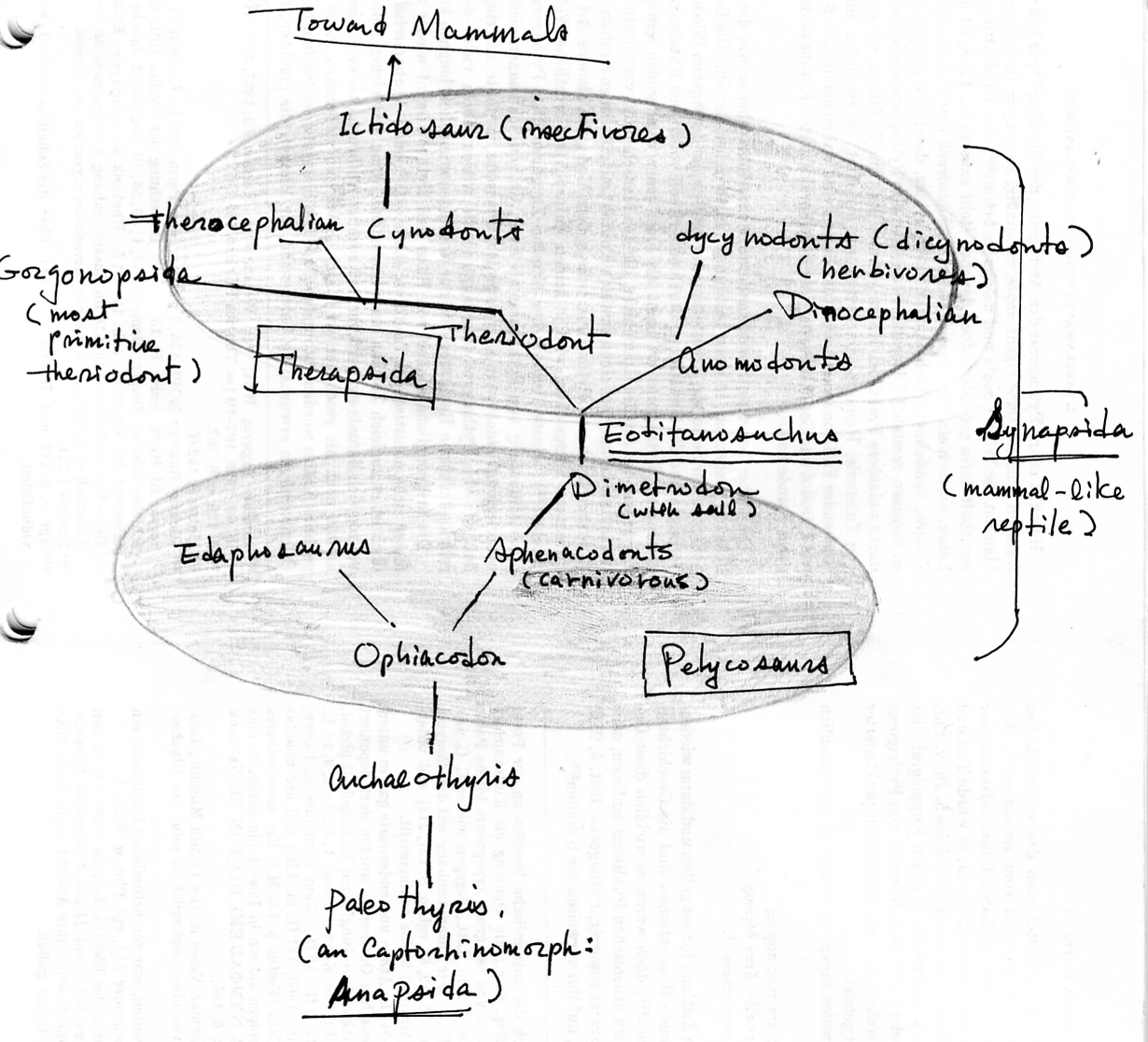
Eotitanosuchus within the evolution of Synapsida Colbert and Morales, ibid., p. 119 Tracing our ancestry beyond our mammalian lineage, we then come to the ancestor we share with all mammals and reptiles, the famous "four-legged fish" Ichthyostega during the late Devonian period. We have by now traced our ancestry back into the ocean. Yes, we came from the water, literally; we came from fishes. The fish that is the common ancestor we share with all mammals, reptiles, and amphibians is Eusthenopteron, a crossopterygian or lobe-fin fish, still during the late Denovian period. It is this fish from which the amphibians -- Ichthyostega first among which -- are descended. It is the ancestor of all landed vertebrates. Tracing our ancestry beyond the stage of fish, we next come to pikaia, the ancestor of all chordates, including all fish. Yes, this is the common ancestor we share with fishes and all landed vertebrates. This is during the Cambrian era. It was only two inches in length, and swam above the sea floor. Going further back in time, we come to choanoflagellate, "most likely ancestors of all animals" (Campbell, Mitchell, and Reece, Biology, p. 193). Yes, this little swimmer is most likely the common ancestor we share with all other members of the animal kingdom. It is a complicated process how this choanoflagellate became animal-like. Around 700 million years ago, some "ancient representatives of [these] choanoflagellates, which are monoflagellated, heterotrophic, aerobic protists" joined "into hollow, spherical arrangements combining cooperative propulsion with cooperative feeding": association. (De Duve, ibid., p. 188) This is the stage recapitulated in embryology as "blastura". The blastura as the colony of choanoflagellates is the protist ancestor of the animal kingdom. Here a first division of labor among single cells (differentiation) occurs between somatic (locomotive and feeding) and reproductive cells. (Campbell et al., p. 371) This is the differentiation between the genetically transmitted germ and mortal soma. "With time, further mutations flattened the sphere into a miniature, double-walled pancake with a back and belly made of different kinds of cells": the thick, ventral (belly) layer with cells for crawling and food-gathering, and the back (dorsal) layer with those for protection and swimming. (De Duve, ibid.) Out of the somatic cells therefore are further differentiated structural cells (skin). "The animal sometimes raised its central part above the sea floor, creating a space that served as a primitive alimentary cavity." (Ibid.) Here the stage of "gastrula" begins. "Like the parent protist, the organism could reproduce sexually under certain conditions, such as excessive crowding. It did so by way of large, nutrient-laden egg cells, which were released after fertilization and developed into copies of the parental organism." (Ibid.) There is now the differentiation between inner and outer layers, and between protective skin, locomotive (both outer layer), feeding/digestive and reproductive functions (both inner layer) among cells. "The main advantage of the layers is to enable further division of labor among the cells... Junctions holding the cells together and enabling the transport of nutrients among them may also have developed by this time." (Campbell et al, ibid.) "This account tells how the basic features of cellular collectivism -- association, differentiation, patterning, and a genomically inscribed body plan -- could have been first realized in the animal kingdom." (De Duve, p. 190) 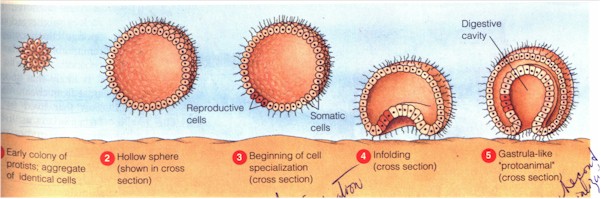
The formation of proto-animal out of choanoflagellates (3) blastura and (5) gastrula The gastrula is diploblast, i.e. two-layered, of which a contemporary representative would be the placozoa, e.g. Trichoplax adhaerens. "Two major lines diverged from the primitive, ancestral diploblast." One is toward sedentarianism. Here the gastrula was re-arranged into a sac-like form, the ancestor of modern sponges, "with an outside layer of flat cells and an inside layer of flagellated collar cells" called chaonocytes which retain the shape of the original choanoflagellates. (Campbell, p. 372; Kristen Bender, General Biology, Laboratory Manual, 2nd ed., p. 191) The flagellating cells lining its interior "fished out bacteria and smaller food particles from the flowing water [which entered the sponge from the top opening and through the pores perforating its body, and] which also provided them with mineral salts and oxygen and carried away waste products." (de Duve, p. 190) Thus this first animal became fixed and "it is only in their immature stages (flagellated larvae) that sponges move about." (Bender, ibid.) The division of labor among its collection of cells arranged in radial symmetry however does not mean that it has any tissues, organs, or organ systems -- these higher level differentiations among cells. (Ibid.) In the other direction the digestive cavity of the gastrula was further accentuated to take "the shape of miniature double-walled pouches that opened to the outside by a narrow orifice" and become thus "a segregated digestive cavity" with the rim of the pouch endowed with food-catching appendages. "The outcome was a tiny, primitive medusa, the common ancestor of hydrae, polyps, sea anemones, jellyfishes, and other related organisms known jointly as coelenterates" (animals having coelom or body-cavity) or grouped into the phylum Cnideria (de Duve, ibid.). These are the full realizations of radially symmetric body-plan (Bauplan) and the last of the diploblastic stage. Especially of note here are further intensified differentiations within the Person, not just among cells, but between "departments" or groups of cells, to give rise to true tissues ("organized groups of similar cells adapted to perform a specific function"; Camp. p. 373), and among cells also to give rise to nerve cells, as yet organized in a de-centralized (and hence primitive) nervous system. "Some cnidarians exist only as medusa" which "move freely about in water", "others (such as hydras) only as polyps", a stationary (thus like sponges) cylindrical body with tentacles, "and still others have both a medusa and a polyp in their life cycles." (Campbell, ibid.) Since polyp is asexual and medusa sexual, and there exists in some an alternation of asexual and sexual generations, we see that a secondary differentiation among the Persons of species -- and so within the differentiated reproductive cells -- has begun but not yet reached completion. (Bender, p. 192) The evolutionary lineages of the Kingdom Animalia (From Campbell, Mitchell, Reece, ibid.) Our next stop is the common ancestor we animals share with all plants and fungi. That's right. Today you might see little commonality between plants and fungi on the one side and animals on the other, but that's only because during the past billion and a half years the three brotherly or sisterly lineages have gone their own way and evolved into such different lives. Their common ancestor is the first eukaryotic organism. The primitive phagocyte is an example; it is without as yet any endosymbiont-derived organelles such as mitochondrion, etc. It lived before the oxygen crisis around 1.5 billion years ago. A good modern reference is Giardia: "about 1/1000 of an inch in size, which makes its volume more than 10,000 times larger than that of an average prokaryote" (de Duve, p. 139). It is when we get to the common ancestor of all life on earth -- from bacteria through plants to animals -- that our lineage starts to blur. Let us get a bit familiar with the history. Firstly, the traditional five kingdom classification of all life on earth, first proposed by Robert Whittaker in 1959, has been supplanted since the 1990s by the three domain system of Carl Woese, et al. The five kingdoms are: Monera comprising the single celled prokaryotes, Protista (roughly single celled eukaryotes), Plantae, Fungi, and Animalia, all three of which are multicelled eukaryotes. This system is still in use, with modifications, in Lynn Margulis et al, Five Kingdoms, today's standard book of biological taxonomy. With sequencing and comparison of ribosomal RNA, however, Woese has shown that a three-domain division underlies the traditional five kingdoms, with (eu-)Bacteria first diverging from Archae, from which eukaryotes emerged later. The basic shape of the new tree of life is given below:
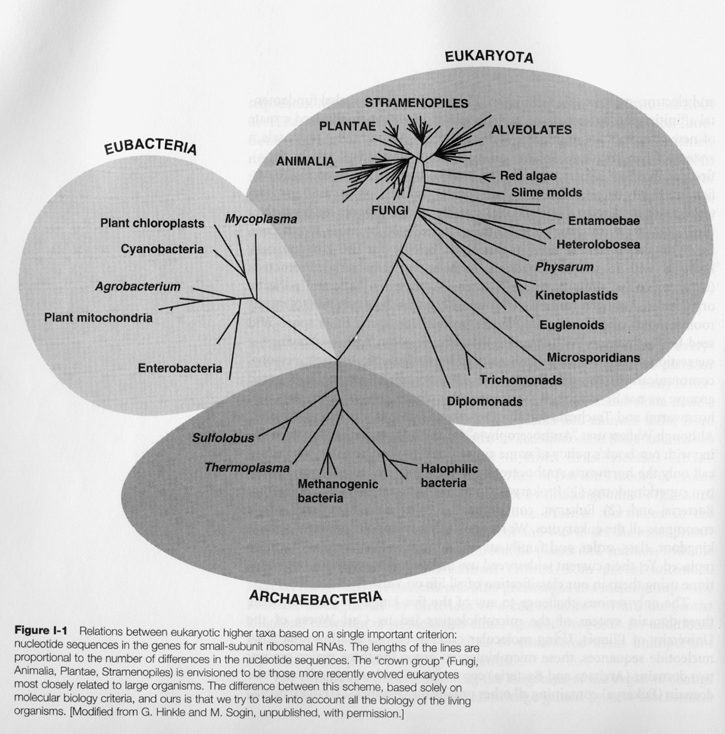
(Taken from Lynn Margulis, ibid., p. 8) The new view has been implying, along with an early origin of life, also an early origin of all three domains, around 3.5 billion years ago. (Roger Lewin, Patterns in Evolution: The New Molecular View, p. 63) More recently, the picture of the evolution of the earliest life has become more complex. The traditional construction of a tree of vertically descending lineages which, when traced backward in time, would converge at a root, has been confounded by the new realization of a large-scale horizontal transfer of genetic materials across the three domains during the earliest times.
Something like the "Bacteria Interaction Sphere." The new picture is like this: 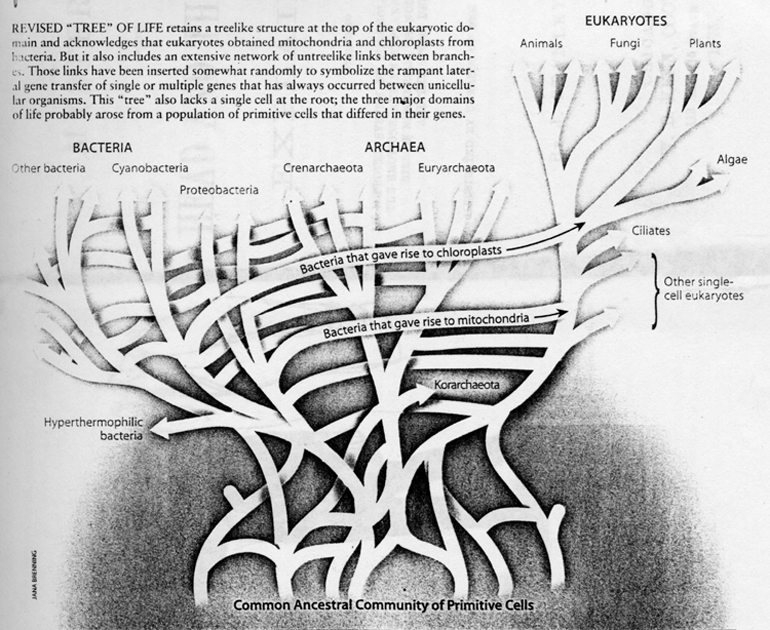
(Taken from Doolittle, ibid.) One must notice here the same pattern we have noted before: From a vast ocean of simple forms only a small portion within (the eukaryotes) complexified into mutlicellularity and emerged to become the most dominant (at least most conspicuous). In the tree diagram, plants and animals, which dominated the activities on the surface of the earth, are mere two small twigs in the corner of Eukarya. It also says about the vast foundation of simplicity that is needed for the evolution therefrom of a thin strand of complexity. More can be said about the universal ancestor of all life:
Within the electron-transport chain (i.e. its mechanism for energy-release) they mostly likely used ferric iron as the final electron acceptor rather than oxygen as do aerobic eukaryotes today. These first organisms were thus chemoautotrophic (de Duve, p. 116).
Within this universal ancestor "pool of life" did one of the three domains emerge first or did they all emerge simultaneously? Evidence seems to point to eubacteria as the most ancient while eukaryote branched off from archaebacteria which itself branched off from eubacteria beforehand.
|
